为便于研究长期发热症状,Petersdorf[1]在1961年通过对100例患者前瞻性观察,首次提出不明原因发热(fever of unknownorigin,FUO)的概念。随着诊断技术的不断发展,不明原因发热的概念也在不断扩展,目前感染性疾病是FUO的最主要原因[2],在20世纪90年代,20%~60%的FUO病属于感染性疾病[3]。而进入21世纪后的10年内,最主要的感染病原为细菌、病毒以及真菌和寄生虫。近年来病毒感染比例呈明显上升趋势。然而传统的分子生物学检测方法需要病原基因组序列的先验知识,因此对于样本中的未知病原需要有新的序列非依赖性的检测手段来处理。
近些年来,随着二代测序技术(NGS)的飞速发展,得益于二代测序通量的高速提升和成本的大幅下降,使得二代测序技术越来越广泛的应用于宏基因组检测未知病原的研究领域中。相对于传统的序列依赖性的核酸检测技术,如PCR技术等,二代测序结合宏基因组技术具有不依赖于病原序列信息的特性,因此对临床样本的检测使得我们能够得到更加广泛的病毒谱,进而也可以发现一些用传统方法无法观察到的病原组成情况[4]。
本研究中,对10份不明原因发热症状的血液样本进行宏基因组测序(metagenomics shotgun sequencing, MSS)时,发现了大量的人细环病毒(torque teno Virus,TTV)相关序列。对这些序列进行组装拼接并分析其进化树后,发现其分属于α TTV、β TTV和γ TTV三个属。
壹
材料与方法
1.1 实验材料与方法
选取10份汉坦病毒IgG抗体检测(ELISA)阴性的不明原因发热的临床血液样本(均为出现发热症状后一至两周内采集),采用Qiagen公司的RNease Mini Plus Kit试剂盒进行核酸提取。病毒基因组的反转录采用Invitrogen公司的Superscript Ⅲ试剂盒进行一链合成,随后加入Klenow片段37 ℃孵育2 h进行二链合成。反转后的cDNA采用REPLI-g WTA Single Cell Kit(Qiagen)试剂盒进行基于多重链置换反应(multiple displacement amplification,MDA)的预富集。采用Thermalfisher公司的Ion Torrent Hi-Q试剂盒构建测序文库。以上步骤均按照相关厂商提供的标准流程进行。测序反应在Ion Torrent PGM(Thermalfisher)平台上完成。
1.2 数据分析
所有原始数据均为从Ion Torrent平台下载的bam格式下机数据,应用本地分析软件Virus IdentificationPipeline(VIP)进行分析[5]。
贰
结果
2.1 测序结果
在10份样本中均发现了不同程度的TTV感染,其中2号和6号样本为单一性的α TTV感染;1号、5号和9号样本为α TTV和γ TTV共感染;7号样本为α TTV和β TTV共感染;3号、4号、8号和10号样本为α TTV、β TTV和γ TTV共感染。α TTV、β TTV和γ TTV的感染率分别为100%、50%和70%(表1)。在这10份样本中,α TTV、β TTV和γ TTV3种TTV基因组覆盖度达到70%以上的概率分别为:60%、30%和40%(表1)。其中8号样本病毒载量最高,α TTV、β TTV和γ TTV3种TTV基因组覆盖度均达到99%以上,αTTV和γTTV的平均测序深度达到了400×以上(表1)。所有匹配读长(Reads hit)在整个病毒基因组上的位置如图1所示。

图1 8号样本中三种TTV(A:TTV, B:TTMV, C:TTMDV)一致性序列的进化分析(红色圆点代表组装出的一致性序列,蓝色方块代表亲缘关系最近的代表株)
Fig.1 Phylogenetic analysis of TTV in 3 genera (A: TTV, B:TTMV, C:TTMDV) in sample 8 (red dotrepresent the assembled consensus sequence, blue square represent the closestreference)
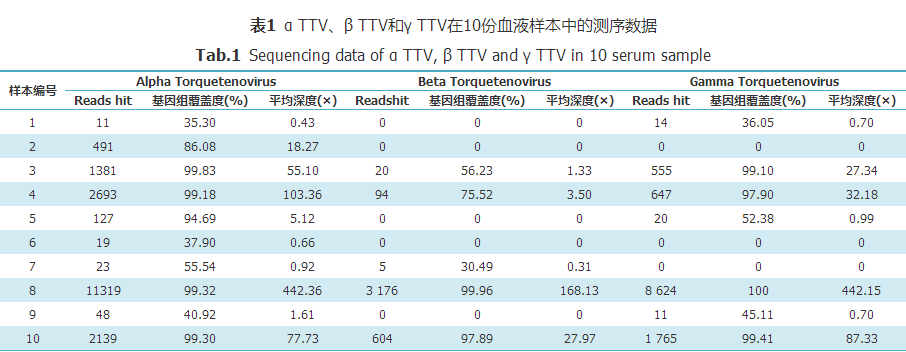
表1 α TTV、β TTV和γ TTV在10份血液样本中的测序数据
Tab.1 Sequencing data ofα TTV, β TTV and γ TTV in 10 serum sample
2.2 进化分析
用10份样本中的测序reads组装出的置信序列与从GenBank引用的TTV各型别的代表株以NJ法构建序列进化发生树。其中8号样本的三种TTV序列进化树分别见图1。
叁
讨论
不明原因发热(FUO)是常见的临床综合征,病因广泛,有报道超过200种疾病能够引起FUO。据统计,在1952-1994年间的FUO病例中,感染性疾病(占比28%)为主要病因,非感染性炎症占比21%,恶性肿瘤占比17%,病因不明占比19%[2]。在非细菌感染样本中,病毒感染呈明显上升趋势。但在无先验经验的情况下,传统免疫学方法和分子生物学手段难以对样本中的未知病毒病原进行鉴定。随着高通量测序技术的发展,越来越多的新的病毒病原被发现。而这些新发现的病毒飞速的革新着我们关于病毒复杂程度的认知。越来越多的证据表明,这些新发现的病毒比传统已知的病毒病原在临床样本中分布更为普遍。传统意义上的病毒组(virome)的概念现在看来仅仅是构成人体宿主宏微生物组的一部分[6]。尽管肠道微生物组的研究是当今的热门,但是血液病毒组对于宿主免疫反应和器官移植的安全依然有着极其重要的指示功能[7][8]。
近些年来,关于环状、复制起始蛋白编码的单链DNA病毒[circular,replication initiatorprotein(Rep)encoding,single-strand DNA virus,CRESS-DNA virus]的报道越来越多。这些病毒曾经被认为只能感染植物或动物,但近10年的研究发现,该类病毒具有十分宽广的宿主范围,包括人在内的脊椎动物到无脊椎动物均包含在内[9]。这类病毒包括:Anellovirus;Circovirus;Cyclovirus;Gemycircularvirus;Gyrovirus;Smacovirus等[10,11,12,13]。这些病毒的病原学特性依然不清楚。而有报道称Anellovirus能占到整个病毒组的70%左右[14]。在本研究中,核酸提取虽然采用的是RNA提取试剂盒,但也能够采集到足量的病毒DNA,满足检测的需求。在本研究及类似的研究中[15],作为DNA病毒的TTV的核酸均由RNA提取试剂盒所提取。
人细环病毒(torque teno virus,TTV)最早于1997年在日本一例输血后发生急性感染的非甲-戊型肝炎患者血清中发现[16],被怀疑与肝炎相关。随后TTMV(Torque teno mini virus,)与TTMDV(Torque teno midi virus)分别于2000年和2007年[17]相继被发现。随后的研究表明,人TTV在世界范围内存在广泛的分布[18,19]。国际病毒分类委员会(International Committee on Taxonomy of Viruses,ICTV)将TTV归类于一个新建立的科:指环病毒科(Anelloviridae),目前该病毒科共包含9个病毒属:α TTV(Alpha torquevirus)、β TTV(Betatorquevirus)、γ TTV(Gamma torquevirus)、δ TTV(Delta torquevirus)、ε TTV(Epsilontorquevirus)、η TTV(Eta torquevirus)、ι TTV(Iota torquevirus)、θ TTV(Thetatorquevirus)、ζ TTV(Zeta torquevirus)。目前国际病毒分类委员会主要以ORF1基因为基础对TTV进行分类。以碱基序列35%的一致性为阈值来划分以人为宿主的3个属:Alphatorquevirus genus(Torque teno virus,TTV),Betatorquevirus genus(Torque tenomini virus,TTMV)和Gammatorquevirus genus(Torqueteno midi virus,TTMDV)[15]。除此之外还有其他以各种动物为宿主的属:猪细环病毒(Torque teno sus virus, TTsuV),猫细环病毒(Torqueteno felis virus),狗细环病毒(Torque teno canis virus)以及树鼬细环病毒(Torque teno tupaia virus)[20],近些年来,在啮齿类以及蝙蝠中也发现了相关的TTV。到目前为止,TTV被认为分为5个主要的进化簇,包含数10株准种。各准种间在ORF1上的碱基序列的差异超过35%[21]。在本研究中,二代测序产生的读长在α TTV、β TTV和γ TTV3个属中均有分布。其中α TTV感染率最高,在10份样本中均有检出,γ TTV和β TTV排在其后,分别在7份和5份样本中检出。全部样本中未发现其他属TTV相关的测序读长。而全部的10份样本中,除1号样本外,其余9份样本的α TTV检出水平高于β TTV和γTTV。在β TTV和γ TTV之间,除了2号和6号样本两者均未检出,7号样本β TTV检出高于γ TTV以外,其余7份样本中γ TTV检出均高于β TTV。
TTV基因组为单股负链环状DNA,无包膜。基因组大小从2.86 ~2.91 kb(TTMV)、3.24~2.35 kb(TTMDV)到3.6~3.9kb(TTV)不等,包含4个开放阅读框(openreading frame,ORF),约2.6 kb,以及一个非编码区(untranslated region, UTR),约1.2 kb。不同TTV分离株之间编码蛋白的氨基酸差异可以高达47%~70%[19,22]。这种差异性分布并不均一,在编码区明显多于非编码区。
对于TTV基因组如此高变的解释是TTV拥有非常快的突变速率[22],但有趣的是,作为一个DNA病毒、在缺乏自我复制机制、必须利用宿主具有高校准能力的DNA聚合酶系的情况下,依然拥有如此高的突变速率。合理的解释是由于其单链的基因组结构和编码复制过程中涉及的蛋白的能力所致[23]。另一种解释是仅有小部分TTV拥有完整的感染宿主细胞的原件,而大多数情况下感染宿主细胞是1个多准种的协同过程。也有报告称Epstein-Barr virus(EBV)在TTV的感染过程中起到了协助的作用[24]。
TTV在世界范围内广泛存在,但分布并不均一,报道过的在人群中的分布最低到5%[18],最高达到90%[25]。与第一次被发现相似,TTV普遍存在于各类血液疾病、器官移植、肿瘤甚至牙周炎患者的体内。在健康人群中同样存在广泛的分布[25],并且这种分布随年龄的增加而提高[26]。
TTV具有作为免疫标志物的巨大潜力,其广泛的分布意味着TTV已经建立起与宿主之间成功的互作关系。在一些采用PCR方法对TTV的病毒血症进行长期监控的研究中[27],即便TTV阴性患者在免疫抑制状态下也会变为阳性。提示病毒在组织中比在外周血中有更高的免疫耐受。Kincaid等[28]的研究发现,一种由TTV编码的miRNA所介导的免疫逃逸机制,加上TTV在正常人群中的广泛而均一的分布,使得TTV成为了一种免疫低下或免疫缺陷的标记。这种miRNA靶向作用于N-myc (and STAT)的反应原件(N-myc interactor,NMI),由此产生对干扰素的干扰作用,并促使细胞增殖裂解。
由于缺乏合适的细胞系,TTV难于进行体外培养。同时缺乏相关的血清学方法来验证病毒代谢产物与特异性的抗病毒免疫反应。而TTV本身基因组高度的变异以及与宿主相互作用的复杂性,都使得TTV潜在的病原特性难以评估。因此,迄今为止尚未有明确的研究证据证明人细环病毒与任何临床症状具有直接的关系。在本研究中,未检测到明确致病病原可能由以下原因导至:一是在越来越多的报道中所高度怀疑的TTV潜在的病原特性[15];二是由于10份血液样本均为出现发热症状7~14d内所采集,而真正的致病病原的基因组序列信息有可能因为宿主已处于非急性期而含量偏低,导至在宿主血液中已经难以检测到,推测宿主免疫系统对发热致病病原产生应急反应和免疫应答后,宿主免疫水平进入相对的受抑制或低下状态。而在采样时,宿主可能依然处于免疫受抑制或免疫水平低下状态,从而导至TTV在血液中的检出率升高。虽然TTV的病原学特性并不明确,这种现象提示TTV、宿主与致病病原三者间存在一定的相关性。另外,考虑到患者并未发现恶性肿瘤或非感染性炎症等症状,那么其他不明病因导至的发热,以及由此而来的宿主免疫水平的下降,则可能导至潜伏的TTV在血液中的检出率升高。
选自中华实验和临床病毒学杂志, 2018,32(2)